




研究内容 Research
線虫 C. elegans は土壌に生息する非寄生性の動物で,成虫の体長が1mm程度しかなく,大腸菌を餌に実験室で簡単に飼育することができます.また線虫は体や卵殻が透明なため,受精や初期胚発生の様子を生きた個体内で観察できること,変異体の取得など遺伝学的解析が容易であることなどから,発生や神経科学の分野などで広くモデル生物として用いられています.私たちは線虫の生殖腺を用いた解析から,受精直後にはオートファジーやエンドサイトーシスといったリソソーム分解系が活性化し,膜成分の積極的な分解を行っていることを見出してきました.配偶子から接合子への転換期には細胞の膜成分も積極的な分解を介して再編成されていると考えられます.
図1 リソソーム分解系の活性化による母性・父性由来成分の分解
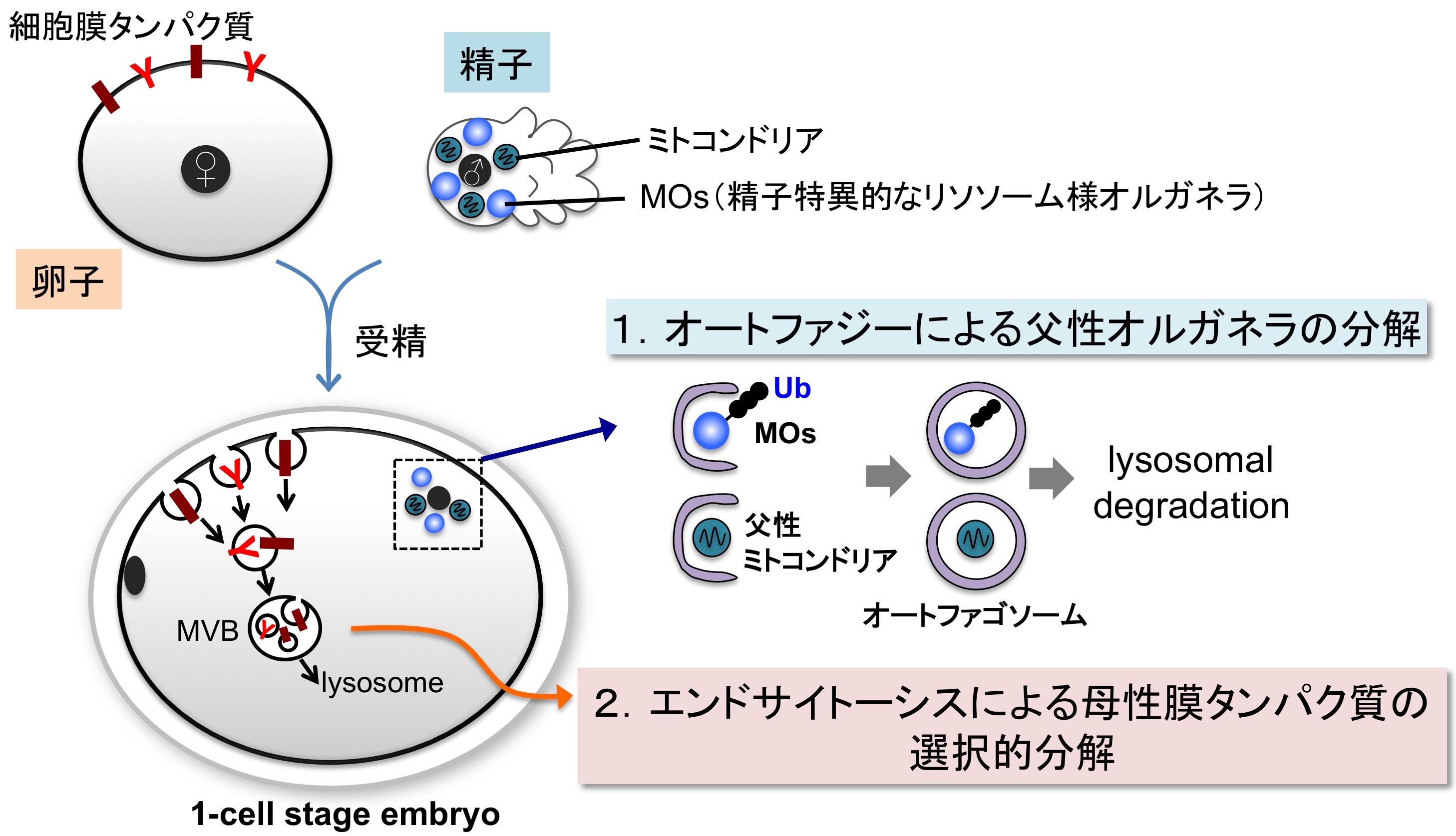
受精後に活性化されるリソソーム分解系. 受精直後にはオートファジーとエンドサイトーシスが一過的に活性化され,それぞれ特異的な膜成分の分解を行なっている.
1. オートファジーによる父性ミトコンドリアの分解
オートファジーは細胞質の成分(タンパク質やオルガネラ)を二重膜で囲い込んでオートファゴソームを形成し,リソソームと融合することで内容物を分解する大規模分解システムです.私たちは線虫受精卵において,受精によって持ち込まれた精子由来ミトコンドリアとそこに含まれるミトコンドリアDNAがオートファジーによって選択的に捕捉・分解される現象を見出しました.また,この分解はミトコンドリアDNAが母方からのみ伝わる“母性遺伝”の仕組みでもありました.現在はどのようにして精子由来ミトコンドリアのみが選択的に認識されオートファゴソーム膜にリクルートされるのかに注目し,そこに関わる因子の探索を行っています.また,このオートファジーによる精子ミトコンドリアの分解の生理的・進化的意義の理解も目指しています.
図2 オートファジーによる精子由来ミトコンドリアの選択的分解
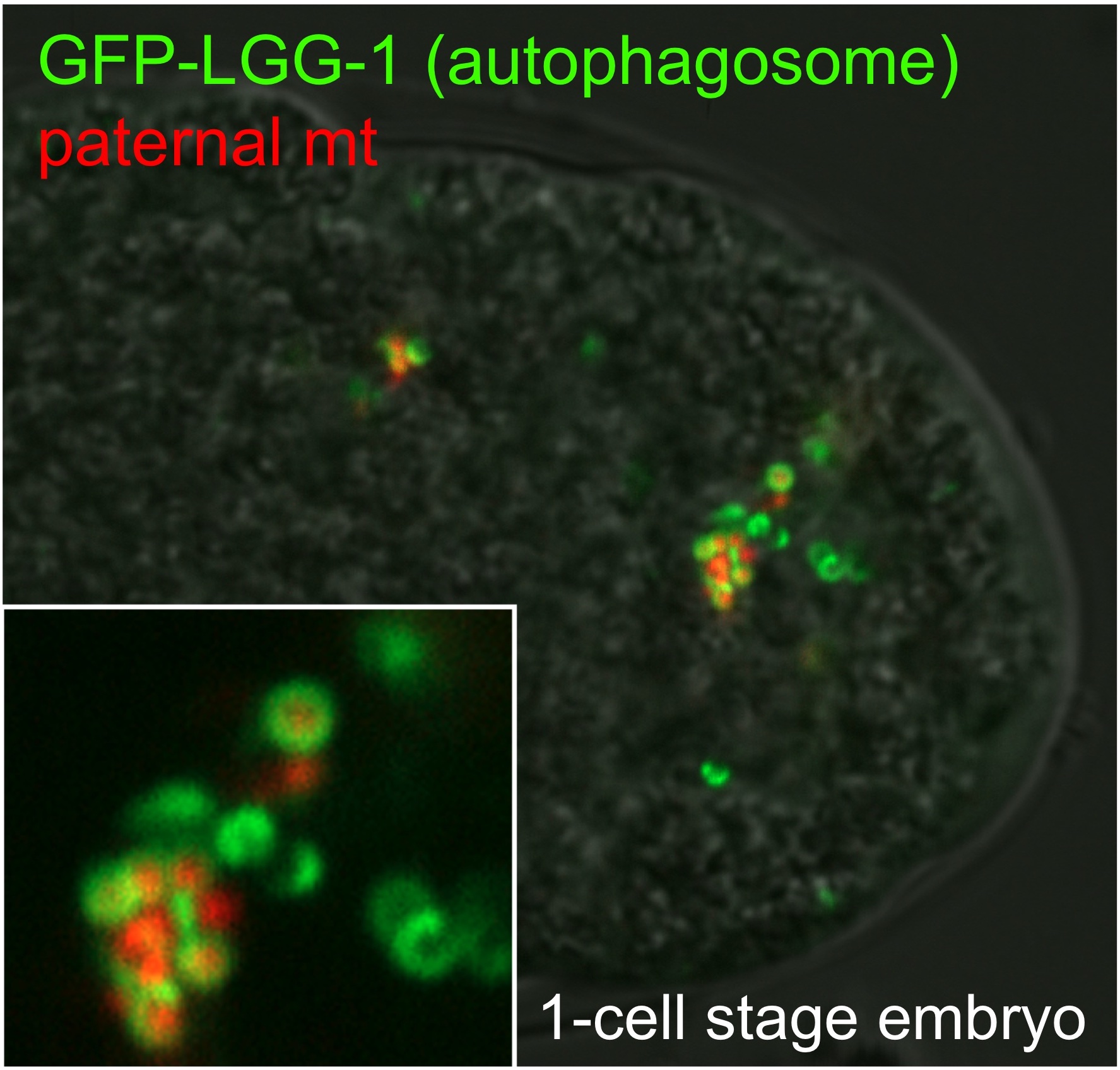
オートファジーによる精子由来ミトコンドリアの選択的分解. 侵入した精子ミトコンドリア周囲にオートファゴソームが膜が形成される様子を生きた受精卵で観察した.
2. 受精後に誘導されるエンドサイトーシスによる細胞膜成分の分解
エンドサイトーシスは環境からの栄養素やシグナル因子の取り込みを行うメカニズムであるとともに,細胞膜上の受容体の量を調節することで,細胞外からのシグナル伝達の強度やタイミングも制御しています.私たちは線虫受精卵ではエンドサイトーシスが一過的に活性化し,卵子に由来する一群の細胞膜タンパク質が積極的に分解されていることを見出しました.また,この分解には基質タンパク質のユビキチン化が関与していることも明らかにしてきました.現在は特異的ユビキチン化のメカニズムを探索するとともに,受精のシグナルがどのようにしてユビキチン化経路を活性化するのか,そのシグナリングのメカニズムについても解析を進めています.
図3 K63結合ユビキチン化を介したエンドサイトーシスの制御
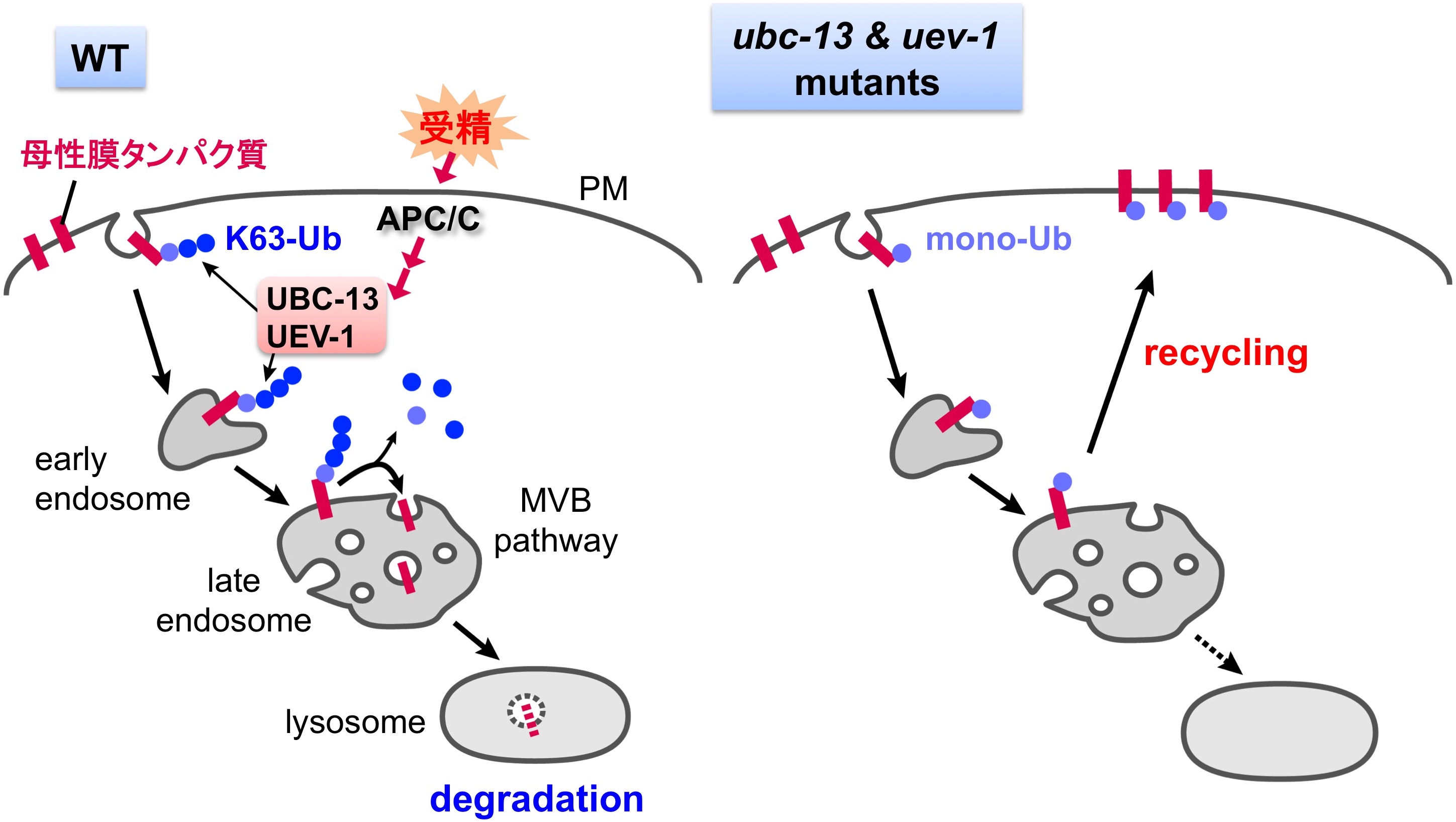
エンドサイトーシスにおけるユビキチン化の関与. 受精後にはK63結合ユビキチン化が誘導され,卵子に由来する母性膜タンパク質の分解を制御している.